
|Articles|September 11, 2017
Reactivating cones possibility in addressing RP
Glucose entrapment in the retinal pigment epithelium is responsible for central vision loss in retinitis pigmentosa. Injecting wild-type rods into the eye to restore glucose transportation or injecting glucose into the subretinal space can facilitate cone regeneration.
Advertisement
Reviewed by Henry J. Kaplan, MD
Take-home: Glucose entrapment in the retinal pigment epithelium is responsible for central vision loss in retinitis pigmentosa. Injecting wild-type rods into the eye to restore glucose transportation or injecting glucose into the subretinal space can facilitate cone regeneration.
Dr. KaplanReactivating lost cone photoreceptor vision might be a breakthrough in addressing retinitis pigmentosa (RP). This important finding is based on the results observed in an autosomal dominant pig model of RP (P23H), the most common autosomal-dominant mutation in RP in the United States. The key to the breakthrough was restoration of glucose trapped in the retinal pigment epithelium (RPE) that ultimately reactivated the dormant cones.
“Patients with RP go blind regardless of the gene mutation ultimately because of loss of cone vision resulting from cone dormancy,” Henry Kaplan, MD, reported.
While most RP mutations occur in rod-specific photoreceptor genes, there is progressive rod destruction, loss of peripheral vision, and dark adaptation. The subsequent loss of cone function, or cone dormancy, results in functional blindness.
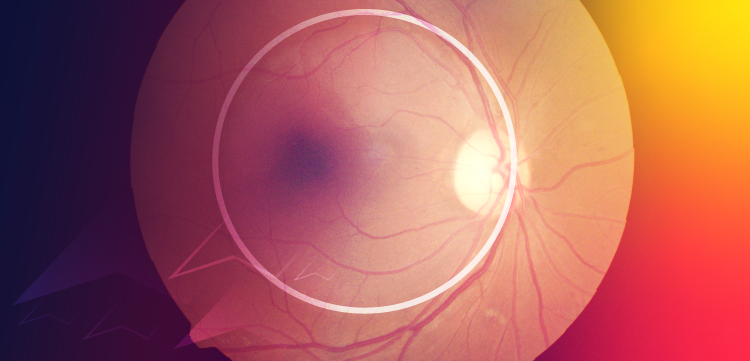
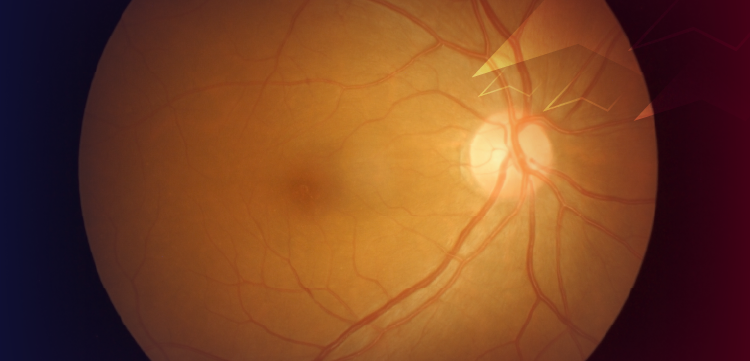
Cone dormancy in RP results in viable cone nuclei for many years after outer segment/inner segment degeneration. (A) Glucose transport to the subretinal extracellular space and cone inner segment is essential to provide energy for regeneration of cone outer segment. (B) Degeneration of rod outer segment and loss of contact with apical microvilli results in entrapment of glucose in the RPE. Ensuing glucose starvation of cone photoreceptors results in outer segment degeneration (C) and inner segment disassembly but with maintenance of viable cone nuclei (D). Courtesy of Henry Kaplan, MD.
This important functional loss (i.e., cone dormancy) is defined as the deterioration of central vision that follows rod degeneration in RP because of cone degeneration, Dr. Kaplan explained. He is the Evans Professor of Ophthalmology, chairman, Department of Ophthalmology and Visual Sciences, and director, Kentucky Lions Eye Center, University of Louisville, Louisville, KY.
Stages of cone degeneration
Two sequential stages occur in cone degeneration that culminate in visual loss–first, the degeneration of the cone outer segments, and second, the disassembly of the cone inner segments.
Most interestingly, however, is that despite this two-stage malfunction, the cones remain viable for a time. “They can remain viable for months or years despite that the cone inner and outer segments have deteriorated,” he emphasized.
The primary culprit in cone degeneration is glucose starvation because the glucose becomes entrapped in the RPE, causing the cones to lose their outer segments and possibly the inner segments, thus leaving the nucleus behind.
“Loss of these functional structures diminishes cone-dependent, high-resolution central vision, which is utilized for most daily tasks,” according to Dr. Kaplan.
Porcine model
The porcine model used in the study of cone dormancy was characterized by progressive loss of cone function from birth, so Dr. Kaplan and colleagues were able to see by electrophysiologic evaluation the gradual deterioration of visual function over time to the point that only the nuclei of the cells remained.
Fourteen days after birth, the transgenic pig retina was essentially normal with no dramatic differences and the outer nuclear layer was intact. By 30 days, however, a dramatic loss of rod photoreceptors was observed.
By 60 days a thin layer of cells was present; and by 120 days only nuclei of the cone photoreceptors are left, with no inner or outer segments present. In this model, Dr. Kaplan pointed out that the scotopic response was extinguished and the photopic response, which represents the cones, began to decline at the 60-day time point.
Because cone survival depends on viable rods, the question arose regarding the ability of transplanted, rod-induced pluripotent stem cells, or rod precursor cells, to recover the function of cones and reverse the damage in the cone inner and outer segments.
“Following rod stem cell transplant, we observed reassembly of the cone inner segments, regeneration of cone outer segments, and increased electrophysiologic function within 1,000 µm from the transplant margin for at least 3 months after the transplantation in all directions,” Dr. Kaplan reported. “Unexpectedly, cone inner segment reassembly was present even beyond outer segment regeneration, suggesting a different mechanism for this effect.”
Glucose metabolic changes
Another interesting observation was that glucose metabolic changes occurred in the regenerating cones following stem cell transplantation. “We observed that the transplanted rod stem cells induced the expression of many glucose metabolic enzymes necessary for cell membrane synthesis of cone outer segments,” Dr. Kaplan said.
This finding led to the observation that within 72 hours of direct injection of glucose into the subretinal space, cone outer segments regenerated and restored retinal electrophysiologic function.
“We concluded that glucose starvation was the cause of cone outer segment degeneration,” Dr. Kaplan said.
These results were also confirmed in a mouse model of RP.
“By transplanting wild-type rods to restore glucose transport, or by directly replacing glucose in the subretinal space to bypass its retention in the RPE, we can regenerate cone functional structures, reactivating the dormant cells,” the investigators reported in Cell Reports (Wang et al. 2016;12:372-85).
Henry J. Kaplan, MD
E:
This article was adapted from a presentation that Dr. Kaplan delivered at 2016 Precision Ophthalmology meeting. Dr. Kaplan has no financial interest in any aspect of this report.
Advertisement
Related Content
Advertisement



Latest CME















Advertisement
Advertisement
Trending on Modern Retina
1
Retina World Congress 2026: Identifying real-world durability gaps in wet AMD treatment
2
Retina World Congress 2026: The case for earlier surgical intervention in diabetic macular edema
3
Retina World Congress 2026: Diagnosing macular telangiectasia in a field of mimickers
4
Retina World Congress 2026: Rethinking macular hole surgery without gas tamponade
5